I am interested in a broad diversity of topics relating to the study of animal behavior (ethology), including animal communication, neuroethology, behavioral ecology, evolution of behavior, comparative cognition, and animal welfare. Although much of our group’s current research focuses on lacertid lizards (family Lacertidae), we occasionally work with other reptiles, invertebrates (beetles), and canids (wolves and dogs). Our work involves observational and experimental studies of animal behavior in both the laboratory and the field. Regarding communication my interest lies in the design, information content and evolution of animal signals. Our theoretical contributions include a revised definition of communication, a defense of the central role of information in animal communication, and an integrative conceptual framework for the study of the evolution of signal unreliability and deceit. Our empirical work has emphasized chemical and, more recently, visual communication in lizards. Lizard visual signals include both dynamic, movement-based displays (foot shakes, head bobs and tail displays) and color patterns. Foot shakes have been described in several agamid, iguanid and teiid species but they are almost completely unstudied in lacertid lizards. We have found that many lacertids possess a complex repertoire including 3-4 stereotyped foot shake displays that differ in their structure and are performed in different contexts, presumably making available to receivers different types of information. At least in some species, a particular type of foot shake display is elicited by an approaching predator and has a pursuit-deterrent function.
Regarding communication my interest lies in the design, information content and evolution of animal signals. Our theoretical contributions include a revised definition of communication, a defense of the central role of information in animal communication, and an integrative conceptual framework for the study of the evolution of signal unreliability and deceit. Our empirical work has emphasized chemical and, more recently, visual communication in lizards. Lizard visual signals include both dynamic, movement-based displays (foot shakes, head bobs and tail displays) and color patterns. Foot shakes have been described in several agamid, iguanid and teiid species but they are almost completely unstudied in lacertid lizards. We have found that many lacertids possess a complex repertoire including 3-4 stereotyped foot shake displays that differ in their structure and are performed in different contexts, presumably making available to receivers different types of information. At least in some species, a particular type of foot shake display is elicited by an approaching predator and has a pursuit-deterrent function. Currently, our research addresses the study of coloration and chromatic (color) signals in lacertid lizards. Animal colors have evolved to serve different adaptive functions, such as camouflage, thermoregulation, warning or startling predators, and communicating with conspecifics. Recent research has stressed the complexity of color traits, which are often the result of multiple, often conflicting, selection pressures. For example, selection for effective communication with mates and rivals favors flashy, highly conspicuous coloration, but conspicuousness also makes color signals more detectable to unintended receivers (e.g. predators, parasitoids). Many lacertids solve this trade-off by allocating conspicuous color patches to the lateral and ventral body surfaces, where they are less visible to predators. Using reflectance spectrophotometry we have found that some of the conspicuous color patches used for social communication include an ultraviolet (UV) component which is invisible to humans but adds to the complexity of the signal as perceived by other lizards. In some species, chromatic variables of the UV-reflecting color patches are correlated with bite force and body condition and may thus act as reliable signals of individual quality, particularly in males.
Currently, our research addresses the study of coloration and chromatic (color) signals in lacertid lizards. Animal colors have evolved to serve different adaptive functions, such as camouflage, thermoregulation, warning or startling predators, and communicating with conspecifics. Recent research has stressed the complexity of color traits, which are often the result of multiple, often conflicting, selection pressures. For example, selection for effective communication with mates and rivals favors flashy, highly conspicuous coloration, but conspicuousness also makes color signals more detectable to unintended receivers (e.g. predators, parasitoids). Many lacertids solve this trade-off by allocating conspicuous color patches to the lateral and ventral body surfaces, where they are less visible to predators. Using reflectance spectrophotometry we have found that some of the conspicuous color patches used for social communication include an ultraviolet (UV) component which is invisible to humans but adds to the complexity of the signal as perceived by other lizards. In some species, chromatic variables of the UV-reflecting color patches are correlated with bite force and body condition and may thus act as reliable signals of individual quality, particularly in males.
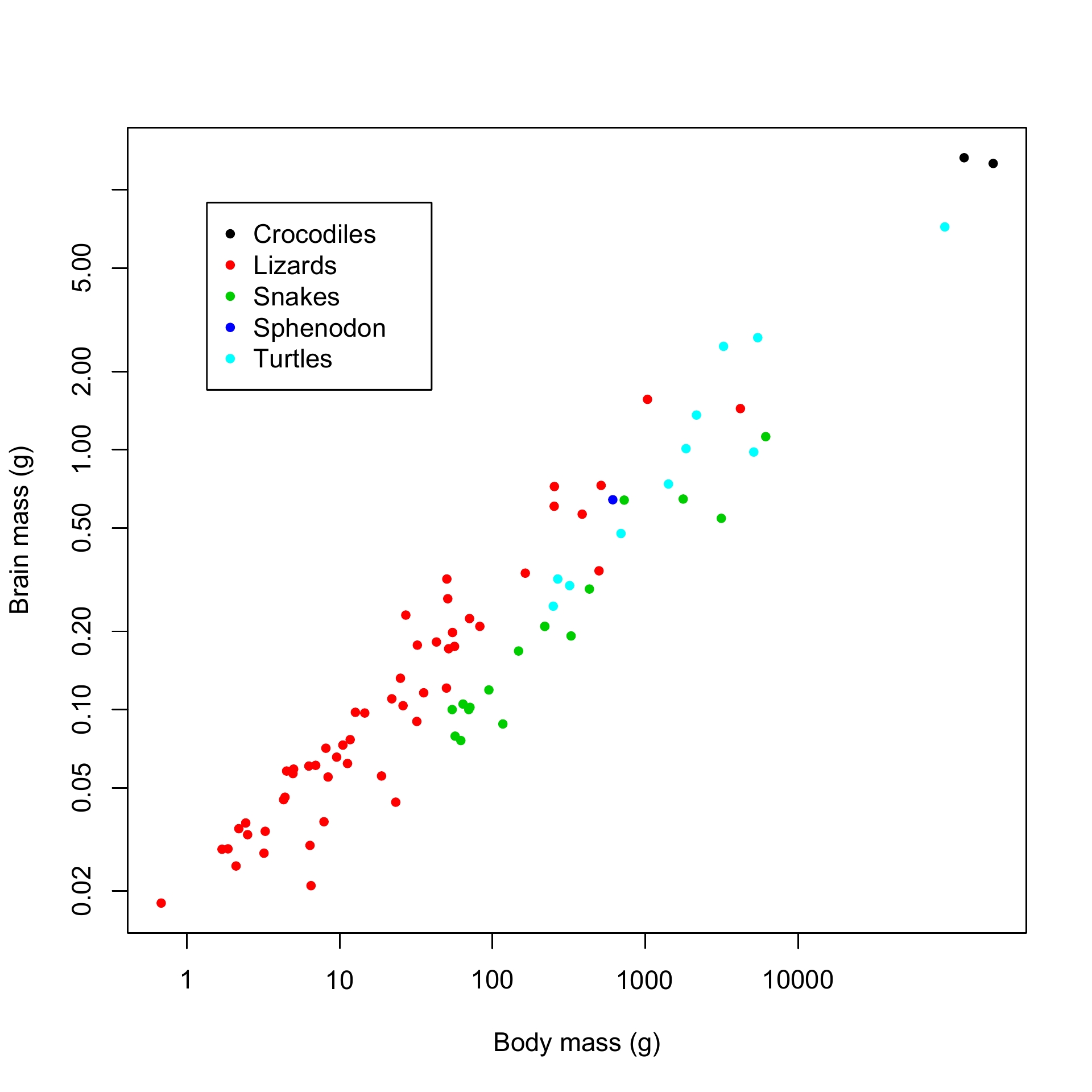
I am also interested in brain size evolution in vertebrates and in the relationship between brain size and cognition. Studies several decades old claiming that reptiles have small brains and are either primitive, stupid, or both are still authoritatively cited. However, a reanalysis of the available data suggests that the differences with other vertebrate classes (birds and mammals), both in terms of brain size/complexity and cognitive abilities, have been grossly exaggerated.